抗体の仕組みと種類を理解しよう
まずは抗体について理解しましょう
免疫化学を活用した抗体技術は、1970年代初期に免疫標識の研究ツールとして用いられて以降大きく進歩し、ライフサイエンス研究の多くの分野において必要不可欠なツールとなっています。免疫化学の基本原理は「特異的な抗体が、特定の抗原と結合し、ユニークな組み合わせの抗原抗体複合体を形成する」ことです。
この記事では、免疫化学および抗体技術を理解するために重要な要素のひとつ「抗体」について、解説します。
抗体とは「感受性のある動物において、抗体の産生を引き起こした抗原と、特異的に結合することが可能な免疫グロブリン」と定義されています。抗体は、外来性の分子による体内への侵入に反応して産生されます。抗体はAbと略され、通常は免疫グロブリン(略称Ig)と呼ばれます。ヒト免疫グロブリンは、構造的および機能的に類似した糖タンパク質(タンパク質82〜96%、糖質4〜18%)のグループで液性免疫に寄与しています。
抗体の構造
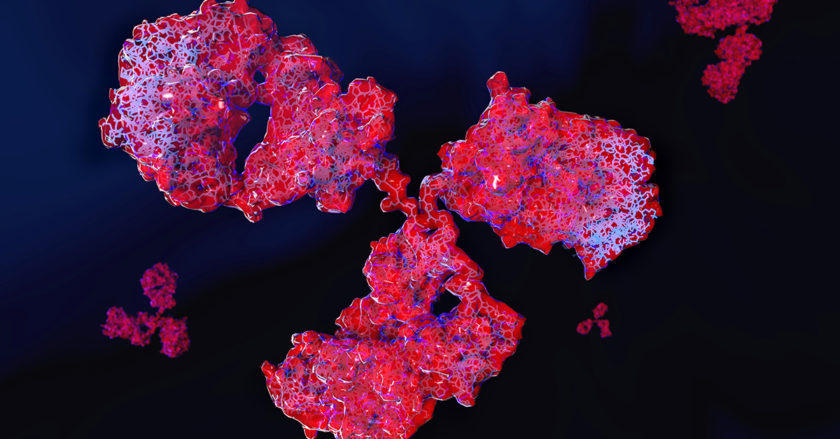
抗体は、4本のポリペプチド鎖からなるY字型のユニットで、1つ以上のコピーとして存在しています。それぞれのY字には同一の重鎖が2本、同一の軽鎖が2本あります。重鎖と軽鎖という名称は、それぞれの相対的な分子量によって命名されたものです。
抗体の構造の図
このY字型のユニットは、F(ab)「アーム」という抗原への結合に重要な、可変性で抗原特異的な2本の部位と、Fc「テイル」という免疫細胞のFc受容体に結合する、定常性の領域で構成されています。Fc「テイル」は、ほとんどの免疫化学手順において抗体を操作するときに有用な「ハンドル」として用いられます。抗体のF(ab)領域の数は、抗体のサブクラス(下表参照)と対応しており、抗体の結合価(抗原に結合可能な抗体の「アーム」の数)を決定しています。
免疫グロブリンのサブクラス

これら3つの領域は、タンパク質分解酵素 パパインによって2つのF(ab)フラグメントと1つのFcフラグメントに切断されます。一方、ペプシンによって切断すると、ヒンジ領域で結合した1つのF(ab’)2フラグメントと1つのFcフラグメントに分かれます。F(ab)フラグメントは抗原を沈降させずFc領域がないことから、in vivo試験では免疫細胞によって結合されません。したがって、酵素消化によるIgG抗体の断片化は、非特異的結合の回避に有用な場合があります。
サブクラスとは
抗体は、Y字型ユニットの数や重鎖の種類に基づき、IgG、IgM、IgA、IgD、IgEの5つのクラスに分けられています。IgG、IgM、IgA、IgD、およびIgEの重鎖は、それぞれγ、μ、α、δ、およびεと記されます。抗体の軽鎖は、ポリペプチド構造のわずかな差異によって、カッパ(Κ)またはラムダ(λ)型のいずれかに分類することができますが、抗体のサブクラスは重鎖によって決まります。
抗体のサブクラスによって、ジスルフィド結合の数やヒンジ領域の長さが異なります。免疫化学法において最も一般的に用いられている抗体は、IgGクラスの抗体です。これはIgGが血清中に放出される主要な免疫グロブリンであるためです。
免疫グロブリンのサブクラス:IgA
血中では単量体として低レベルで存在しています。粘膜表面で最も活性が高く、二量体として存在し、粘膜表面の一次防御を担っています。粘膜内層では、その他のすべての種類の抗体を合わせたよりも多くのIgAが産生されています。
IgAの主な機能は、中和抗体としての作用です。唾液、涙、母乳には、IgAが高レベルで認められます。ヒトでは2種類のIgAのサブタイプが知られていますが、マウスでは1種類しか報告されていません。IgA1は、血清中の総IgAの最大85%を占めていると考えられています。選択的IgA欠損症は、易感染性を増大させる最も一般的な免疫不全疾患の1つです。IgA欠損症は、自己免疫疾患やアレルギー疾患を有する患者によく認められます。
半減期は約5日です。
免疫グロブリンのサブクラス:IgD
2つのエピトープ結合部位を有する単量体の抗体であり、ほとんどのBリンパ球の表面に存在します。
IgDの正確な機能については未だに議論されていますが、B細胞の活性化に必要な抗原受容体として作用するのではないかと考えられています。IgDが好塩基球や肥満細胞に結合してそれらを活性化させ、抗菌因子の産生を誘導することも報告されています。また、自己反応性の自己抗体を産生するBリンパ球の除去においても役割を担っていると考えられています。
IgDは血清中で少量認められる分泌型としても産生されており、分泌型は、δクラスの2本の重鎖と2本の軽鎖からなっています。
IgDの半減期は約3日です。
免疫グロブリンのサブクラス:IgE
このグループの抗体は、粘膜表面、血液、および組織において機能しています。IgEは、2本の重鎖(ε鎖)と2本の軽鎖からなる単量体として存在しています。ε鎖には、4つの免疫グロブリン様定常ドメインがあります。血清中には低濃度で存在し、血清中のすべての抗体の約0.002%しかありません。ほとんどのIgEは、Fc領域を介して、肥満細胞や好塩基球上のIgE受容体に堅固に結合しています。
IgEは過敏症反応において重要な役割を担っており、IgEの産生はサイトカインによって厳密に制御されています。
IgEの半減期は約2日です。
免疫グロブリンのサブクラス:IgG
IgGは、血中で最も豊富に認められる抗体のクラスで、血清中のすべての抗体の最大80%を占めています。IgGは単量体として存在しています。存在量によって4種類のIgGのサブクラスがあり(IgG1>IgG2>IgG3>IgG4)、どのサブクラスが産生されるかは存在するサイトカインの種類によります。
IgG1およびIgG3は、食細胞のFc受容体に高い親和性を示しますが、IgG2のFc受容体への親和性は非常に低く、IgG4の親和性は中程度です。IgGは、循環器系から出て組織に侵入することができます。IgG1、IgG3、およびIgG4は胎盤関門を通過することができ、新生児の保護に役割を果たしています。IgGは補体系の活性化に効果があり、食細胞のFc受容体を用いたオプソニン化に非常に有効です。IgGはFc領域を介してナチュラルキラー細胞にも結合することができ、抗体依存性の細胞毒性に寄与します。
IgGの半減期はサブクラスによって異なり、7〜23日にわたります。
免疫グロブリンのサブクラス:IgM
このクラスの免疫グロブリンは、感染に反応して産生される最初の免疫グロブリンであり、B細胞の膜上や、形質細胞によって分泌される5つのサブユニットからなる高分子として存在しています。IgMは、新生児が最初に合成する免疫グロブリンでもあります。細胞膜結合型IgMと分泌型ではFc領域が異なっています。膜結合型IgMは、Fc受容体にではなく、内在性膜タンパク質として直接的に細胞膜上に存在します。分泌型IgMは五量体分子であり、複数の免疫グロブリンがジスルフィド結合によって共有結合的に接合しています。この構造によって複数の結合部位が供されています。各単量体は、2本の軽鎖(κまたはλのいずれか)と2本の重鎖によって形成されています。五量体であるため、IgMは補体の活性化や凝集の誘発に特に適しています。
IgMの半減期は約5日です。
以上、抗体技術の基本原理を理解するために重要な「抗体」とそのサブクラスについて解説しました。技術を適切に利用し、試行錯誤するためには、原理をきちんと把握することが大切です。抗体を使用した実験を始める前には一度、抗体と抗原について復習してみましょう。
抗体がよくわかる!「抗体ガイドブック 3版」。
この記事以外にも抗体そのものの理論や選び方が分かる記事あります。
理論
- 【研究ツールとしての抗体技術】抗原とエピトープ
- 抗体の仕組みと種類を理解しよう
- 抗原と抗体の相互作用とは【抗体技術の基本原理】
- モノクローナル抗体とポリクローナル抗体の作製と特徴
- 用途に合わせて使い分け。抗体のフォーマットと精製方法
実用
これら抗体の理論や実用、応用、プロトコールの例を掲載した「抗体ガイドブック 3版」をご用意しています。こちらのページで印刷版の請求またはPDFのダウンロードが可能です。


下記フォームでは、M-hub(エムハブ)に対してのご意見、今後読んでみたい記事等のご要望を受け付けています。
メルクの各種キャンペーン、製品サポート、ご注文等に関するお問い合わせは下記リンク先にてお願いします。
*入力必須