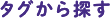基本を学ぶ。酵素と阻害剤の反応速度論

酵素阻害剤について理解するために
酵素阻害剤は酵素による生化学的な反応を阻害するため、研究や医療など様々な用途で活用されています。酵素阻害剤について理解し、利用するには、酵素反応速度論についての基本的な知識が必要です。
この記事では、酵素と阻害剤の反応速度論における基本用語について説明し、ミカエリス・メンテン速度式に用いられるVmaxとKmについて解説します。
知っておきたい酵素阻害剤の基本用語
まずは酵素阻害剤を利用する上で知っておくべき基本用語を確認しましょう。
Km:所定の条件下において、最大反応速度の半分の速度が認められる基質濃度[S]です(図1)。基質と酵素の結合強度とは逆の尺度で、Kmが低いほど親和性が高く、最大反応速度への到達に必要な基質濃度が低いことを意味します。Km値は、pH、温度、その他の反応条件に依存しています。
Vmax:所定の条件下における反応の最大速度(図1)。Vmaxは、すべての酵素部位が基質で飽和したときに達する理論上の最大速度です。これは基質濃度[S]がKmよりも高く、[S]/([S]+Km)が1に近づくときに起こります。
EC50:最大作用(阻害または刺激作用)の50%の作用を生じるために必要な薬物濃度として報告される、薬物の臨床的有効性。阻害反応の場合は、ベースラインと、一定期間にわたり阻害分子に曝露した後の最大反応の間の中間値を指します。この用語は、通常医薬品について用いられます。
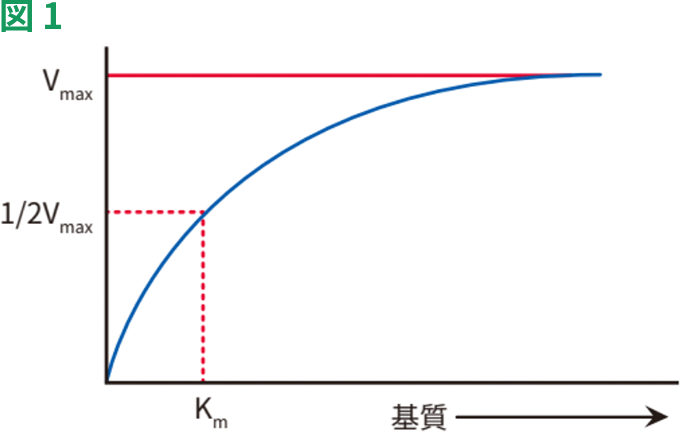
IC50:50%阻害に必要な濃度(図2)。必要な阻害剤の量は、基質濃度、標的への到達性、細胞透過性、インキュベーション時間、使用した細胞型などの様々な要因に依存します。したがって初期濃度の決定には、文献の調査が重要です。公表されているKiまたはIC50値が既知の場合は、酵素活性を最大限阻害するため、これらの値の5~10倍高い値を用います。KiまたはIC50が不明な場合は、幅広い濃度の阻害剤を試し、ミカエリス・メンテン速度式(下記参照)により、Ki値を推定してください。高濃度の阻害剤を用いた場合、阻害がまったく認められないか、逆の反応が認められるということも稀ではありません。(例:プロゾーン効果)
阻害剤の濃度とIC50の関連性
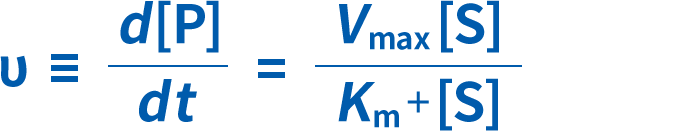
ミカエリス・メンテン速度式
Ki(阻害定数):50%阻害が認められる阻害剤の濃度。ChengおよびPrusoff(1973)*は、IC50が既知である場合に、Kiの算出を単純化する式を開発しました。
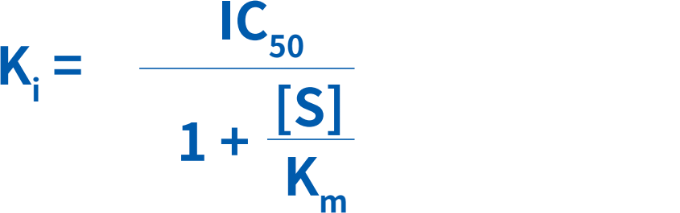
[S]は基質濃度、Kmは反応速度が最大反応速度の半分となる基質濃度(阻害剤の非存在下)です。特定の基質の阻害(一定のKm)に対する阻害剤のKiは一定です。基質によって、KmもKiも異なります。(*Cheng Y. and Prusoff W. H. (1973) Biochem. Pharmacol. 22: 3099-3108.)
ED50:50%の個体において規定量の作用が認められる有効量(濃度ではない)の中央値です。期待される薬物作用の妥当な尺度ではありますが、処方される用量と同等とは限りません。
IC50 は、阻害剤の機能的強度であり、親和性の指標ではありません。化合物の IC50値は、実験条件によって、実験間で異なる場合があります。一方、Kiは絶対値で、阻害剤の結合親和性を反映しています。
KmおよびVmaxの重要性
ミカエリス・メンテン定数であるKmは先述のように、最大反応速度の半分の速度が認められる基質濃度として定義されています。その値は酵素によって著しく異なり、同一の酵素でも基質によって異なります。基質濃度がKm値に等しいとき、酵素の活性部位の半分が基質分子によって占有されています。
Kmについては、次の4点を押さえておきましょう。
- Kmは、単位M(mol/L)の定数です。
- Kmは、速度定数から導かれます。
- Kmは、真のミカエリス・メンテン条件下では、酵素と基質の解離定数の推定値です。
- Kmが小さいと結合が強力であり、大きい場合は結合が弱いことを意味します。
酵素の動態に関するミカエリス・メンテンモデルでは、酵素はまず基質と反応して酵素-基質複合体を形成し、この複合体が分解して、生成物と遊離した酵素になると仮定されています。このため、Kmは3つの速度定数で表すことができます(k1、k-1およびk2)。
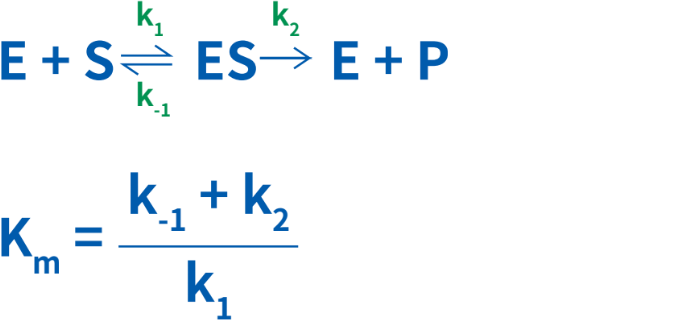
Kmは、温度、基質の性質、反応液のpH、イオン強度、およびその他の反応条件にも依存しています。したがって、酵素-基質反応の特性解析をする際には、定められた特定の条件下で行うことが重要です。Km値の変化は、反応液中に活性化因子または阻害剤のいずれかが存在することを示しています。
基質濃度が非常に低い場合、反応の初速度は基質濃度[S]に比例し、1次反応となります。[S]が高くなるにつれ、反応の初速度の上昇率が低下し、[S]に比例しなくなります。このような条件下では、反応は混合された反応になります。さらに[S]が高くなるにつれ、反応速度は[S]に非依存的になり、漸近的に一定速度に近づきます。この時点で、反応は0次反応となり、酵素は基質で飽和したとみなされます。酵素-基質複合体および生成物が形成される経過は、図3に示すとおりです。
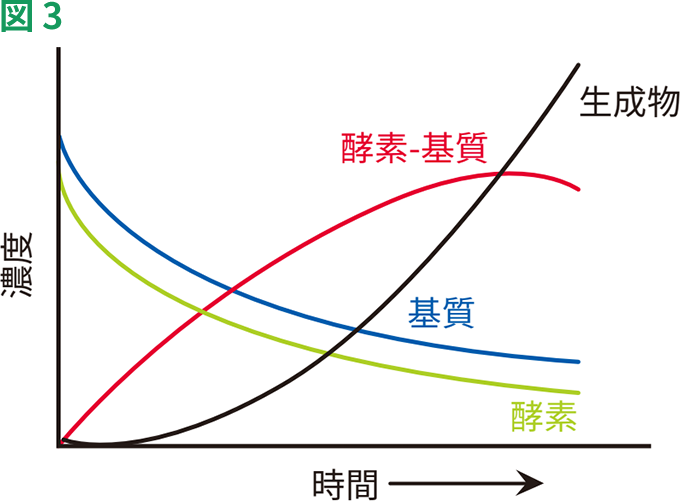
時間依存的な酵素反応の構成要素の相対濃度
Vmaxは、最大反応速度であり、すべての酵素が酵素-基質複合体として存在するときに達する速度です。Vmaxは、所定の条件下において到達可能な最大反応速度を表しています。
Vmaxについては、次の4点を押さえておきましょう。
- Vmaxは、単位s-1の定数です。
- Vmaxは、理論的な最大反応速度で、実際に到達することはありません。
- Vmaxに到達するには、すべての酵素分子が基質と結合する必要があります。
- [S]が高くなるにつれ、漸近的にVmaxに近づきます。
初期酵素濃度が既知であれば、Vmaxからk2の値を推定することができます。k2は1次速度定数であるため単位時間で表されます(毎分または毎秒)。これは代謝回転数または触媒定数kcatとしても知られています。代謝回転数は、酵素が基質で完全に飽和している条件下において、単位時間あたりに生成物に変換される基質分子の数です。
飽和基質条件下([S]がKmよりも大きい場合)で反応速度を測定することにより、代謝回転数を容易に推定することができます。一般的に、生理条件下では、[S]/Kmは1未満です。[S]がKmよりも大きい場合、反応の初速度(V0)は次のように表すことができます。

この式において、kcat/Kmは触媒効率の尺度で、この値が大きいほど、より多くの生成物が形成されます。
ミカエリス・メンテン速度式が適用できない阻害
ミカエリス・メンテン速度式は、不可逆的阻害には適用することができません。これは、阻害剤が酵素と強力な共有結合を形成し、離れることがないためです。したがって、不可逆的阻害剤の有効性は、結合が生じる速度によって判定されます。非常に一般的な不可逆的阻害の例は、アセチルコリンエステラーゼ(AChE)とジイソプロピルフルオロリン酸(DFP)の反応です。DFPは、AChEの活性部位におけるセリン残基のヒドロキシル基と共有結合を形成します。形成された複合体は非常に安定しているため、新しい酵素が合成されてはじめて、正常な神経機能が回復します。
アロステリックに制御される酵素も、ミカエリス・メンテン式には適合しません。アロステリック酵素における基質濃度と最大初速度の関係式は、双曲線ではなく、S字曲線になります。これらの酵素は複数の結合部位を有しており、阻害剤または活性化因子の結合によって活性が制御されます。
KmおよびVmaxの判定方法①Lineweaver-Burkプロット
KmおよびVmaxは、酵素を様々な濃度の基質とインキュベートすることにより、実験的に推定することができます。結果は、基質濃度[S]に対する反応速度(V)のグラフとしてプロットされ、これによって双曲線が得られます。反応速度とKmの関係は、次の式で表すことができます。
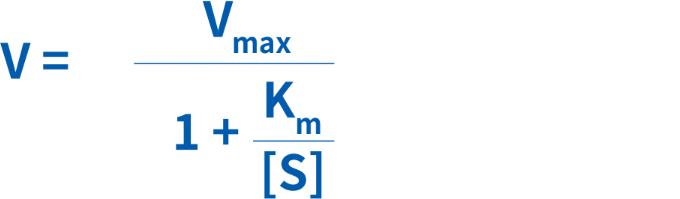
専門家でも、すべての実験データに対して最適な双曲線を適合させ、Vmaxを正確に推定することは困難です。より正確に実験データに曲線や直線を適合させ、VmaxおよびKmを推定するため、ミカエリス・メンテン式の変形方法が開発されています。ただし、どの方法にも利点や欠点があります。
Lineweaver-Burk二重逆数プロット(図4)は、もっとも一般的な方法の1つで、ミカエリス・メンテン式を次のように改変します。
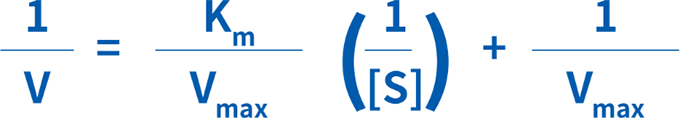
1/[S]に対して1/Vをプロットすると直線が得られ、Y切片=1/Vmax、傾き=Km/Vmax、X切片=-1/Kmとなります。Lineweaver-Burkプロットは、データの直線化にもっとも広く使用されており、もっとも正確なKmおよびVmaxの推定値が得られます。しかし、この方法では、低い基質濃度(すなわち1/[S]および1/Vの最高値)において得られたデータに、過度な重みが加えられてしまいます。
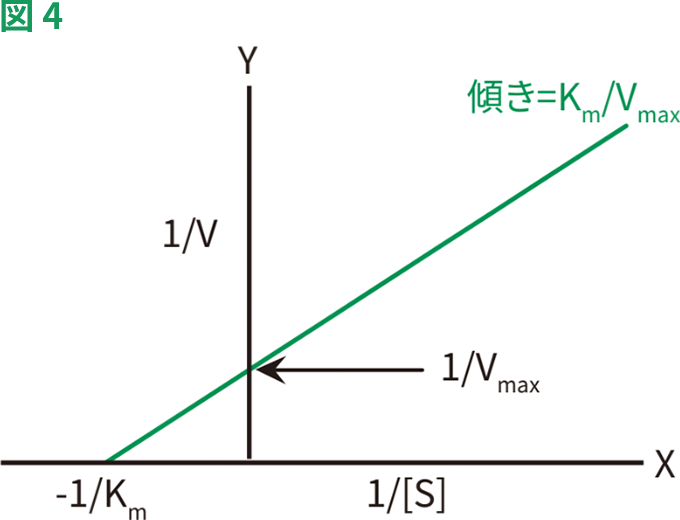
典型的なLineweaver-Burkプロット
KmおよびVmaxの判定方法②Eadie-Hofsteeプロット
酵素の動態を図的に表す別の方法に、Eadie-Hofsteeプロットがあります(図5)。このプロットでは、ミカエリス・メンテン式が次のように改変され、反応速度と基質濃度の比の関数として反応速度がプロットされます。
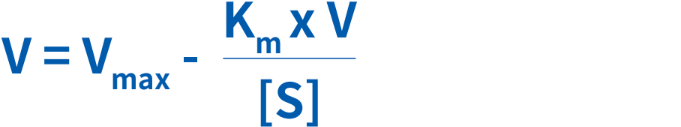
反応速度Vを、V/[S]に対してプロットすると、直線が得られ、Y切片がVmax、X切片がVmax/Km、傾きが–Kmとなります。この方法では、Lineweaver-Burkプロットとは異なり、あらゆる基質濃度範囲においてすべてのデータポイントに等しい重みづけがされます。この方法の欠点は、X軸とY軸のいずれも独立変数ではなく、両方が反応速度に依存していることです。実験や計測器の誤りが、両軸に大きく影響します。
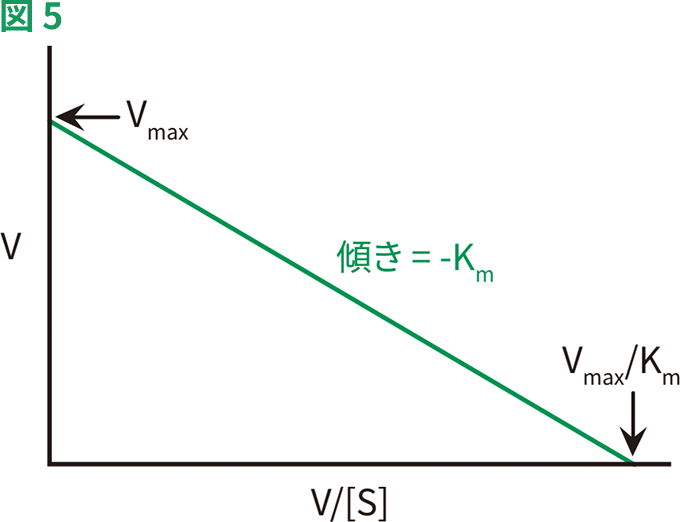
典型的なEadie-Hofsteeプロット
触媒効率
ある基質に対して作用する異なる酵素の反応速度、または異なる基質に作用する同一の酵素の反応速度を比較するとき、相対的な触媒効率を算出すると、特定の条件下において、どの酵素がどの基質にもっとも適しているのかを知ることができます。
酵素が定常状態で作用する場合、考慮すべき酵素の動態パラメータは、kcat(基質から生成物への変換の触媒定数)およびKm(ミカエリス・メンテン定数)です。ここでは、kcatは代謝回転数であり、特定の時間内にどれくらいの基質が生成物に変換されるのかを示しています。kcat/Km比は、触媒効率と等しく、酵素の比較に用いられます。この効率の尺度は、速度を制限しているのが生成物の産生なのか、反応液中の基質の量なのかを判定する際に有用です。一般的にkcat/Kmの上限は、酵素の活性部位への基質の拡散速度によって決まります。
一定の条件下では、ある酵素が2種類のやや関連した基質に作用することがあります。この場合、各基質にはそれぞれのKm値があるため、各基質との相対的な反応速度を考慮しなければなりません。しかし、酵素の特異性がKmのみによって決まる場合、[S]/Kmの比が1を超えて大きくなるにつれ、どの基質の方がよいのかを判定するにはkcatが最良のパラメータになります。2種類の基質(xおよびy)が同時に同一の酵素の作用を受けるとき、相対反応速度について次の式を作成することができます。
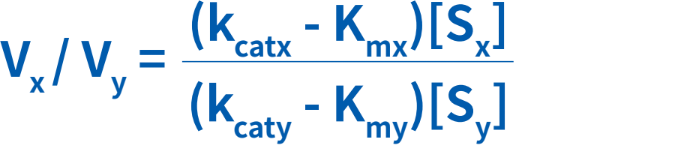
ここでは、VxおよびVyは、基質xおよびyと酵素の反応速度です。2種類の基質の基質濃度は、[Sx]および[Sy]と表現されています。
以上、酵素と阻害剤における反応速度論の基本知識について解説しました。式や記号が多く敬遠されがちな領域ではありますが、阻害剤についてより詳細な理解をするためには、まず基本をきちんと身につけることが肝要です。
<無料PDFダウンロード> 阻害剤 選択ガイド
この阻害剤選択ガイドでは、酵素に対する阻害剤や受容体への阻害剤の作用機序について解説し、適切な阻害剤選びに役立つ情報をご紹介しています。
▼こんな方にオススメ
・最適なプロテインキナーゼ阻害剤を選びたい方
・各種シグナル阻害剤の背景知識を学びたい方
・これから阻害剤を使った実験を行う可能性がある方

下記フォームでは、M-hub(エムハブ)に対してのご意見、今後読んでみたい記事等のご要望を受け付けています。
メルクの各種キャンペーン、製品サポート、ご注文等に関するお問い合わせは下記リンク先にてお願いします。
*入力必須